2021年第6期评论
基于二氧化碳的生物制造:从基础研究到工业应用的挑战
任杰1,曾安平2
(1中国农业科学院植物保护研究所,植物病虫害生物学国家重点实验室,北京 100193;2德国汉堡工业大学,生物过程与系统工程研究所,德国 汉堡 D-21073)
引用本文: 任杰, 曾安平. 基于二氧化碳的生物制造:从基础研究到工业应用的挑战[J]. 合成生物学, 2021, 2(6): 854-862
Citation: REN Jie, ZENG Anping. CO2 based biomanufacturing:from basic research to industrial application[J]. Synthetic Biology Journal, 2021, 2(6): 854-862
DOI: 10.12211/2096-8280.2021-086

识别查看全文
摘 要
在过去的几十年里,人们在二氧化碳(CO2)捕获和利用方面做出了巨大的努力,但是通过生物技术大规模利用CO2还缺乏市场竞争力,迫切需要新的方案和技术。谭天伟和Jens Nielsen的团队(刘子鹤等,2020)最近回顾了第三代(3G)生物炼制技术中生物固定CO2方面的进展和挑战,对原材料、碳固定途径和所涉及的关键因素、能源供应和它们将二氧化碳同化为生物质的效率,以及随后基于3G的产品进行了出色的总结和探讨。该文还介绍了3G生物炼制的前景,指出了存在的挑战,并为未来的发展提供了前瞻性的建议。文章也讨论了整合多种碳固定途径和来自化学、生物和过程工程的技术以实现CO2闭环固定和利用的机会和挑战。除了技术方面,文章还强调有必要进一步增加社会、政治和经济激励措施。本评论简要介绍刘子鹤等综述文章的主要内容,并进一步讨论了基于CO2的生物制造从基础研究到工业应用的几个值得关注的问题:①在原子和分子水平上对碳碳键生成的机理进行更深入的基础和定量研究,以显著提高固碳途径的关键酶和代谢模块的效率;②在代谢途径及细胞水平上,对固碳反应与代谢网络的相互作用进行系统的定量研究;③在生物炼制的意义上,将物理、化学和电化学CO2捕获和转化方法与生物过程相结合,同时考虑产品回收的下游处理;④从工业应用的角度来看,基于自养合成的生物制造存在多个技术瓶颈和经济限制,这些问题短期很难解决,混合营养生物合成(使用CO2和混合碳源)是一个实用的解决方案;⑤大多数CO2固定途径及其产品仍处于“概念验证”阶段,需要更多的工程研究来实现从“0到1”到“1到100”的技术需求,从而真正为碳中和做出贡献。

全 文
本世纪人类面临的最大挑战之一是能源转型和气候变化,这两者都与二氧化碳(CO2)的排放和利用息息相关。迫于大众对频繁极端气候的高度关注和强大舆论压力,世界各主要工业国家都纷纷公布甚至提前原定碳达峰和碳中和的时间点。尤其是中国最近在这方面的重大举措,提出力争于2030年前达到CO2排放峰值,并在2060年前实现碳中和,使得CO2的减排、捕捉和利用成为各领域的重大课题和迫切任务。为实现上述目标,需要工业技术和国家相关产业政策的根本性变革,使能源系统从化石能源转化为以可再生能源为主,制造业从不可再生碳资源转化为以可再生碳资源为主,基于CO2的生物制造从长远来讲是实现这两大目标的最佳途径。
基于CO2的生物制造在文献中也常被称为第三代生物炼制技术,以区别于主要以淀粉及其他含糖物质为原料的第一代生物炼制技术和以木质纤维素等生物质为原料的第二代生物炼制技术,其特征是利用微生物及藻类细胞工厂在光或电等可再生能源的驱动下将CO2等碳一化合物转化为生物能源、化学品及材料等。生物制造原则上具有易于大规模生产、条件温和、选择性好、环境友好等优点,符合绿色生态的理念,近年来基于CO2的生物制造尤其受到广泛的重视。北京化工大学谭天伟教授课题组和Jens Nielsen教授课题组(瑞典/丹麦)合作撰写发表在Nature Catalysis的综述文章“Third-generation biorefineries as the means to produce fuels and chemicals from CO2”(Liu et al, 2020)总结了多年来生物法固定CO2的研究,从原料、固碳途径、能量利用和产品四个大方面系统地介绍了以CO2为基础原料的第三代生物炼制。这篇综述文章内容详实丰富,不但对大量相关文献进行了梳理,指出了CO2基生物制造的难点与重点,并提出了有益的建议和展望。本文简要介绍这篇综述文章,并结合自己的相关研究从基础研究和工程应用的角度提出一些看法和建议,意在抛砖引玉。
Liu等指出,CO2的人为排放量每年高达330亿吨,其来源十分广泛,如工业生产废气、垃圾填埋废气、生物质气化、车辆及呼吸排放等。不同来源的CO2含有不同的杂质,如氮氧化物和硫氧化物,宿主微生物细胞对于杂质的耐受将是影响生物炼制的重要因素之一。又比如,工业烟道气的温度与生物发酵的温度相差极大,对烟道气进行冷却将付出成本,温度耐受也将是考验宿主微生物的指标之一。笔者认为,如何综合利用高温气体的能量为生物合成提供能量甚至作为下游产品回收的能量,值得进一步探索,比如利用温度使CO2与某物质反应生成活性物质参与生物合成并循环利用。理论上,CO2可以和环氧化物在高温下反应生成环碳酸酯,若能通过生物或化学反应使其循环利用,则可以起到降低CO2活化能的效果。此外,可考虑生物法和物理法相结合,物理法中CO2的标准化前处理会提升生物合成的稳定性,而且利用CO2压缩气建立高压CO2生物反应器将有利于相关的酶反应平衡向合成方向移动。
固碳途径方面,Liu等的文章对8条已知CO2生物固碳途径(图1)进行了系统的分析和阐述,包括6条天然固碳途径,即卡尔文循环(Calvin-Benson-Bassham cycle, CBB cycle)、Wood-Ljungdahl途径(亦称为还原乙酰辅酶A途径,reductive acetyl-CoA pathway)、二羧酸/4-羟基丁酸循环[dicarboxylate/4-hydroxybutyrate (DC/HB) cycle]、3-羟基丙酸/4-羟基丁酸循环[3-hydroxypropionate/4-hydroxybutyrate (HP/HB) cycle]、3-羟基丙酸双循环(3-HP bicycle)和还原三羧酸(TCA)途径(reductive TCA cycle),以及2条人工设计并已经证实的固碳途径,即还原甘氨酸途径(reductive glycine pathway, rGlyP)和巴豆酰辅酶A/乙基丙二酰辅酶A/羟基丁酰辅酶A循环[crotonyl-coenzyme A(CoA)/ethylmalonyl-CoA/hydroxybutyryl-CoA, CETCH cycle]。卡尔文循环是植物及藻类固定二氧化碳的天然途径,其关键酶为D-核酮糖-1,5-二磷酸羧化酶/加氧酶
(D-ribulose-1,5-bisphosphate carboxylase/oxygenase,
RubisCO),该酶的催化效率非常低下,每分钟仅可催化几个分子反应,而多年来对于该酶的改造一直都没有很好的成功案例。二羧酸/4-羟基丁酸循环、3-羟基丙酸/4-羟基丁酸循环和3-羟基丙酸双循环天然存在于一些自养微生物中,通过焦磷酸硫胺素(ThDP)活化后的羰基碳或是活泼的羰基α-碳作为受体固定CO2,裂解出乙酰辅酶A或丙酮酸为溢出产物,这些天然途径中固碳反应的能量消耗都比较高,难以实际应用。还原三羧酸途径可以看作是三羧酸循环的逆向反应,可以固定2分子二氧化碳溢出1分子乙酰辅酶A,这一途径可以被高压二氧化碳驱动,有望成为后续研究的重点之一。文章还介绍了3条理论设计的CO2固碳途径,如丙二酰CoA-草酰乙酸-乙醛酸(MOG)途径等,通过两步固碳反应最终溢出产物为乙醛酸;尽管从反应步数和理论能量利用效率上优于上述的天然途径,然而途径中从辅酶A到丙酮酸的固碳反应仍有较高的能量壁垒,是否可以高效地完成反应仍需实验进一步验证。与此相比较,CETCH循环在人工设计时避开了辅酶A固定CO2的反应,利用了如巴豆酰辅酶A羧化/还原酶固定CO2消耗的NADH来降低能量壁垒,使得多步级联反应可以顺利在体外完成,不过其体内可行性尚待进一步证实。与上述通过循环反应固定CO2不同,Wood-Ljungdahl途径和还原甘氨酸途径是通过线性反应固定二氧化碳的,其对中心代谢的依赖或对其的干扰较小,我们认为从实现工业应用所需的高通量角度考虑,这可能是这两条途径的独特优势。总体而言,目前关于二氧化碳固定反应与代谢网络相互作用的研究还很有限,对于其相互作用网络(包括调控网络)的重构及系统定量分析具有重要的科学研究价值,值得关注。

图1 六条天然及两条人工构建(CETCH cycle及rGlyP)的CO2固定途径示意图及主要特征
(除CO2及HCO3−外,甲酸也可看作为CO2固定的分子载体,每条途径的溢出代谢物和关键酶都用颜色突出显示;另外,也给出了每条途径所消耗的ATP与还原当量、涉及的酶数量。还原甘氨酸途径中的GCS系统作为例子展示其核心固碳酶机理的复杂性和深入研究的必要性。MCR—丙二酰辅酶A还原酶;4-BUDH—4-羟基丁酰辅酶A脱水酶;PCS—丙酰辅酶A合成酶;ACLY—ATP-柠檬酸裂解酶;KOGR—2-氧代酮戊二酸合成酶;CCR—巴豆酰辅酶A羧化/还原酶;FDH—甲酸脱氢酶;RuBisCo—核酮糖-1,5-二磷酸羧化酶/加氧酶;GCS—甘氨酸裂解系统;CODH—一氧化碳脱氢酶)
Liu等的文章分析了各条途径的优缺点、能量来源、底物以及产物、关键酶的种类以及酶活等。文章特别指出,固碳途径实际应用中的关键性因素为氧气敏感性、ATP需求、热力学、酶动力学和CO2利用形式。氧气敏感性与ATP需求挂钩并与后续产物需求密切相关,热力学与胞内NAD(P)H/NAD(P)的比例相联系,酶动力学决定着细胞生长的速度,受体是与CO2本身还是其碳酸氢根形式结合影响固碳的效率和条件优化。笔者认为,不同固碳途径主要差异之一在于对ATP的利用效率,二氧化碳被固定后往往得到一个羧基,而对羧基的活化及后续官能团的转化往往都需要消耗ATP。Wood-Ljungdahl途径、还原TCA途径、还原甘氨酸途径和CETCH循环消耗能量比较少,而卡尔文循环、DC/HB循环、HP/HB循环和3-HP双循环都额外消耗了较多的ATP(图1)。如何避免ATP的消耗是构建高效固碳途径的重要因素之一,因此对固碳途径有必要进行更深入、更基础的研究,如核心固碳酶对小分子结合及转运的作用机制,这是提高固碳效率及构建新型途径的关键问题。固碳反应的化学本质是亲核进攻反应,亲核基团携带电子进攻CO2中显电正性的碳原子,形成羧酸或其衍生物。由于CO2中的碳原子已经处于八隅体的稳定结构,具有较强的动力学惰性,因此亲核进攻需要克服较高的活化能才能完成,这就需要亲核试剂本身处于较为活泼的高能态且通过酶与底物的结合降低活化能,因而固碳酶往往都有精细而独特的作用机制。
以还原甘氨酸途径为例,其核心固碳酶是甘氨酸裂解体系(glycine cleavage system, GCS),其可以逆向利用CO2、5,10-亚甲基四氢叶酸和氨(NH3)合成甘氨酸,同时固定2分子一碳化合物(CO2和甲酸)和1分子氨(图1)。甘氨酸裂解体系是由三个酶(P蛋白、T蛋白、L蛋白)和一个载体H蛋白组成的多酶复合体,通过载体H蛋白上硫辛酰赖氨酸臂作为活性亲核基团,接受并转移CO2。还原甘氨酸途径为被认为是固碳途径中最具优势的途径之一,反应步骤较少,ATP需求较低,途径中的酶对氧气的耐受性比较好,对中心代谢干扰小,有较好的工业应用潜力。最近还原甘氨酸途径在E. coli、Saccharomyces cerevisiae、Clostridium drakei等微生物中获得成功表达,使得宿主细胞可以利用甲酸和CO2合成有机酸及氨基酸等初级代谢产物。但是,若单纯以甲酸和CO2作为碳源的话,细胞生长非常缓慢,难以达到对工业微生物宿主的要求。其中一个主要的原因是对固碳核心机制的理解还远远不足:甘氨酸裂解系统中各组分蛋白间的相互作用关系、硫辛酰化载体H蛋白的三种形态(Hox、Hred、Hint)以及其自我保护与解除保护、硫辛酰赖氨酸臂的合成以及携带碳一单位移动的方式(代谢通道)、影响因素,都缺乏系统定量的分析,阻碍了对GCS进行工程化的研究和优化,遑论以合成生物学手段对其进行重新设计和改造。最近,我们对H蛋白的自我保护与解除保护过程进行了分子动态动力学模拟,从原子水平揭示了T蛋白诱导Hint解除保护的过程,找到了氨甲基在释放过程中历经的几个阶段所对应的关键氨基酸残基位点,其突变可以明显提高或降低GCS体系的活力,为rGlyP的合理调控提供了定量的理论基础。文献中对于Wood-Ljungdahl途径的研究已经有很多报道,但其中一氧化碳(CO)的结合及一众辅酶A依赖的固碳途径中CO2的结合同样有待于更深入的定量研究。对其他途径的研究,情况也大体如此。这些生物固碳途径在异源微生物尤其是工业微生物中的表达和应用于生物合成,还处于研究的起步阶段,已显示其困难性和局限性,需要综合结构生物学、计算生物学、机器学习、生物化学及代谢分析等进行合成生物学多学科交叉研究。此外,基于原有CO2固定反应的新型代谢途径改造,也非常具有科学研究和实用价值。如Bouzon等向高丝氨酸代谢途径引入了外源的转氨酶和醛缩酶,使之可以裂解成丙酮酸和甲醛,实现一种新的一碳代谢循环,CO2固定与一碳代谢的相互作用关系也是未来研究值得关注的重点之一。
近日,Science杂志在线刊登了中国科学院天津工业生物技术研究所名为“Cell-free chemo-enzymatic starch synthesis from carbon dioxide”的文章。马延和研究员带领的团队经过6年的努力,终于在体外利用CO2通过化学-酶级联转换的方式成功合成了淀粉,其效率在特定的条件下可以达到植物天然合成淀粉的8.5倍。这一开创性的工作为未来实现无土化的粮食生产提供了理论上的依据,具有重要的学术价值和潜在的巨大社会意义。该工作构建了4个级联转换模块,分别是CO2到甲醛的C1活化模块,甲醛到3-磷酸甘油醛的C3缩合转化模块,3-磷酸甘油醛到6-磷酸葡萄糖的C6模块以及最后的聚合合成淀粉的Cn模块。其固碳的过程为CO2和氢气在金属催化剂的作用下合成甲醇,甲醇经酶催化氧化得到甲醛,3分子甲醛在甲醛裂合酶(formolase, fls)的作用下直接缩合成三碳化合物。催化甲醛缩合的fls是2015年由美国科学家通过计算机理性设计由苯甲醛缩合酶改造而来,因其能够直接缩合3分子甲醛而备受关注,但其活力一直很低,导致甲醛积累产生毒性,严重限制了其进一步应用。马延和团队对fls进行了成功的改造,使其活力提高了4.7倍,大大降低了甲醛的积累,提高了合成通量,突破了C1合成的关键瓶颈。可以预期的是,以该工作构建的合成模块为基础,理论上将会有更多的化合物可以从CO2合成而来,大大推进C1生物合成的发展。在欣喜实现从0到1突破的同时,我们还应该看到未来实现产业化(1到100)过程中的困难和挑战所在。高达14个酶(包括辅因子及副产物循环)参与的多酶体系体外大规模生物合成面临着许多难题,如现有途径中能量ATP的供给来源于多聚磷酸,大规模多聚磷酸再生循环利用还不成熟;酶的用量、稳定性及成本控制,工业化C1合成反应器,产品分离提纯,都是产业化需要解决的难题。
能量利用方面,由于CO2是能源化合物燃烧和氧化代谢的最终产物,其能量是最低的,利用CO2就必须引入外部能源(图2)。Liu等的文章讨论了依据能量来源的三类固碳方式:光自养合成、化学自养合成及电自养合成。光自养合成主要存在于植物、藻类及绿硫细菌等,通过卡尔文循环、3-羟基丙酸双循环和还原TCA途径等途径利用不同能量和数量的光子。近年来,光自养合成被移植到模式微生物如大肠杆菌或酵母中,但仍处于起步阶段。光自养合成的主要限制因素在于光能的收集效率较低和微生物生长对于光密度的要求较高。化学自养合成主要是宿主微生物可以利用给电子的化学物质来获得能量,如氢气、CO、甲酸及金属矿物质等,笔者认为,还原性的金属矿物质虽然可以作为能量供体,但来源范围相对较窄,和CO2的大规模利用不相符。电自养合成则是利用电能直接或间接给宿主细胞提供能量,其中间接方式与化学自养合成相近,利用电能将CO2通过电化学还原为CO或甲酸再被细胞利用。作者认为,随着光伏电池在世界范围内尤其是我国的迅速发展,通过光伏电池将光能转化为电能,之后再进行直接或间接的电自养合成,其整体的能量利用效率是有可能高于传统的光自养合成的,利用氢气、CO、甲酸同时作为碳源和能源进行生物发酵有很大应用潜力,这也是Wood-Ljungdahl途径和还原甘氨酸途径的优势点之一。目前直接利用电能进行生物合成虽然受到很大关注,但其效率极低,机理还不清楚,文献报道的微生物如巴氏梭菌(Clostridium pasteurianum)直接从电极获得电子进行生物合成并无直接证据,很有可能是通过电极表面的电子传递中间体如氢气而造成的假象。从已有文献看来,间接性通过氢气和甲酸可能是更为现实的电化学CO2生物利用途径,尤其甲酸可能是在有氧条件下更具有吸引力的能量载体。文章指出了电自养合成的关键影响因素,包括宿主的选择、能量载体的溶解度和传质速率、电解槽中CO2的浓度以及电极和微生物的兼容性。笔者认为,在进一步的研究中,解析电亲核宿主的电子转移机制及电亲核宿主作为模式底盘细胞的工程化研究尤其值得关注。与光能比较,电可以达到更高的能量密度,用光能发电再用电能发酵是有效提高能量密度的策略。此外,电能还可以转化自风能和核能,我国特色的特高压输电解决了电能的转移问题,使得电可以作为标准高效原料参与固碳途径。
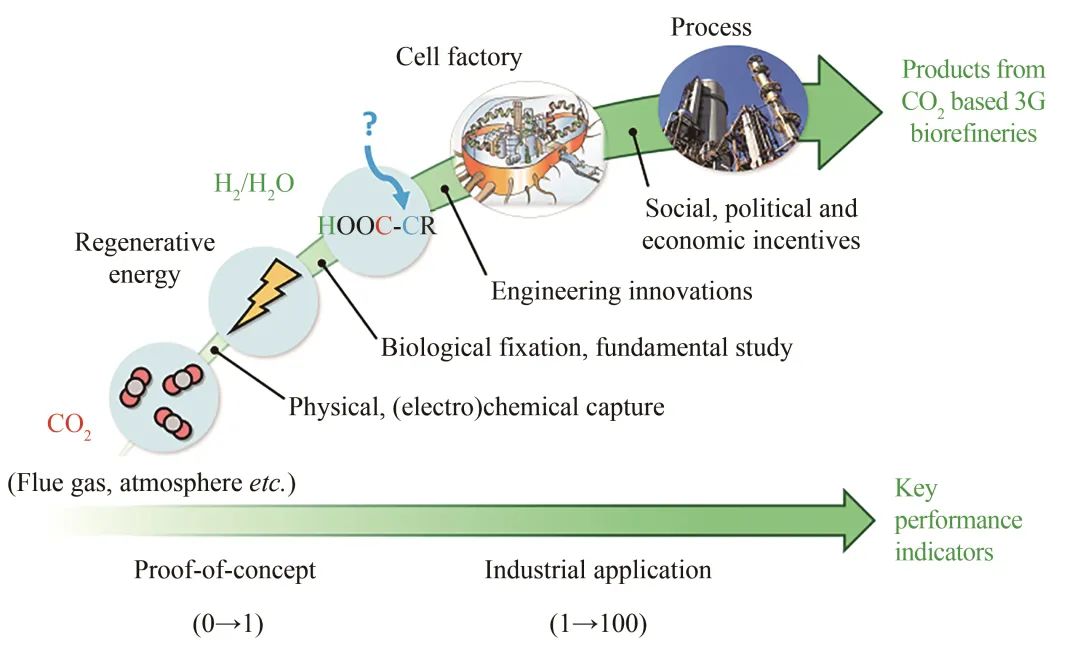
图2 基于CO2的生物制造:从“概念验证”到规模化工业应用
从现实的工业应用角度看来,要解决单纯自养合成中的关键瓶颈,例如底物的传质限制、能量(ATP)及还原当量供应有限,细胞生长慢和产物形成率低,混合营养(mixotrophy)生物合成是有效方案。混合营养涉及共同利用一碳化合物(C1)和有机底物,作为异养和自养生物合成的结合,对其研究还相对较少。Hong和Zeng(2021)介绍和讨论了C1-mixotrophy的不同途径及其潜力,并用示例展示了它们的优势。混合营养生物合成的另一种可能性是提高对天然存在于异养生物中的C1固定回补反应的利用。碳代谢的重新构建可以导致强制C1固定到最终产品中,从而克服异养生物可实现的产品收率的固有限制。从中短期来看,在混合营养中使用C1固定模块的天然或合成途径代表了一种有前途且可行的生物过程策略。为此,需要对C1固定和分解代谢模块的细胞内相互作用进行更多的定量和系统研究。应该更好地研究混合营养中可能的分解代谢物抑制或其他干扰性天然调节机制。逐步改造已建立的生产菌株是提高基于C1原料的生物合成的工业相关性的必要努力。
大规模绿色生物制造的瓶颈在于原材料的可持续性和价格、生产成本以及产品提取方面的制约。实现对以CO2为代表的C1原料的生物利用是解决原料来源的最佳途径和关键技术,合成生物学在提高C1原料转化率,降低成本方面大有作为。目前,第三代生物炼制技术的生产成本总的来说还太高,产品分离提取方面的工作还太少。Liu等的文章最后从产品方面总结目前第三代生物炼制已经可以合成多种燃料和化学品,部分已经开始进入商业化应用阶段,如利用炼钢废气生产燃料乙醇,利用微藻合成脂肪酸类化合物。文章对C1生物合成的最新技术和信息进行了丰富的调查和总结,并对C1生物合成进行了展望,后续值得我们继续思考的有:如引用文献提到用生物质合成氢气并发电,可以将电价降低到2美分/千瓦时,而C1生物合成目标是利用电力和C1化合物合成生物质,很难说这样的电力应用会更有效率,而采用原位(in situ)光或电水制氢和人造光电驱动CO2还原与生物合成相融合的技术研发,有望在这一研究方向做出具有重大突破及影响的工作。
在工业应用方面,目前绝大多数固碳途径和产品还只是停留在“概念验证”阶段,是单一化合物从“0到1”的突破,其技术指标KPI(key performance indicators)远未达到规模化生物制造的需求(图2)。对于大宗化学品的工业化生物制造而言,其产物浓度往往要求在每升上百克(约100 g/L),生产强度在2 g/(L·h)以上。除个别过程及产品外,基于CO2自养型的生物合成通常只能达到mg及g/L的浓度水平,生产强度在g/(L·d)以下。在可预见的将来,要实现从“0到1”到“1到100”的基于CO2的大规模生物制造过程,还需要工程技术的突破与原创的工艺技术相配合。
从工业发展的历史来看,从基础科学到产业的形成,工程技术的突破或原创的工艺技术至关重要。20世纪初,化学工业的革命性成就之一——人工合成氨——就是科学和技术结合的典范。它是由德国卡尔斯鲁厄理工学院的Fritz Haber教授和巴斯夫公司的Karl Bosch博士领导的团队合作完成的。其实,Haber在其工艺原理的研究中所使用的条件[1020 ℃,1 bar (1 bar=105 Pa)]和得到的结果(1904年前后氨的收率约0.005%)是远远无法工业化的,是巴斯夫公司在高压化工过程和设备方面的突破,才催生了世界上第一座高温高压(约450 ℃,300 bar)化学反应器,使得这一过程在1913年得以实现工业化,导致随之的农业革命,为此他们分别于1918年和1931年获得了诺贝尔化学奖。可以说,没有这种科学与工程的紧密结合,很难产生这样划时代的技术!基于CO2的生物合成也需要在生物反应器方面有所创新,与传统生物合成不同,它是体积缩小的反应过程,加之气体溶解度低,高压生物合成具有显而易见的优势,但这方面的研究极少。最近,Steffens等发现在细菌Hippea martima中,仅仅通过提高环境CO2的浓度或分压(40%)即可逆转三羧酸循环,从而实现从CO2和氢气合成丙酮酸。在这一过程中,无需三羧酸循环酶体系以外的酶参与反应,只需其中的柠檬酸合成酶有较高的表达水平。这一现象可能存在于许多别的生物体内,主要是由热力学决定。这为高压CO2生物转化提供了很好的生物学理论基础。
以二氧化碳、甲烷、甲酸和甲醇为代表的一碳化合物(C1)的生物转化及工业应用是合成生物学及绿色生物制造的重大机遇和挑战,是助力碳中和的重大课题。研究重点在于合成生物学、过程系统工程、材料科学及其他基础科学与技术的综合应用,实现一碳生物转化、光电生物技术及工程工艺过程的有机融合,最终实现利用大气CO2和太阳能等绿色能源进行生物制造,为人类缓解气候变化做出贡献。
通讯作者及团队介绍

曾安平博士,西湖大学合成生物学和生物工程讲席教授,校级合成生物学与生物智造中心创始主任。原汉堡工业大学(TUHH)终身教授,生物过程与生物系统工程研究所所长。1990年获布朗瑞克工业大学(TU Braunschweig)生物化工博士学位。之后在德国国家生物技术中心(GBF,现德国国家实验室赫姆霍茨感染研究中心,HZI) 生化工程部作为博士后工作一年即被聘为GBF永久研究员,历任GBF实验室及HZI系统生物学研究组负责人至2006年, 期间在澳大利亚国家科学与工业研究院(CSIRO)从事生物过程开发研究,美国明尼苏达大学化工系访问研究。2005年分别获聘三所德国大学生物化工,生物过程工程及动物细胞系统生物学专业终身正教授,2006年起任职汉堡工业大学。主要研究方向为生物化工/生物工程、系统和合成生物学,电驱动生物合成技术,一碳生物利用及新型蛋白质生物材料,尤其在二元醇及氨基酸生物合成中取得多项成果,实现工业应用。是德国化学工程和生物技术协会专业委员会“生物过程工程”及“系统与合成生物学”专家组成员,曾任“新一代生物制造系统”专业委员会主任、Engineering in Life Sciences主编之一。在国际高水平刊物上发表论文近300篇,编著五本,专利(含申请)20多项,多次成为欧盟、德国科学基金委,联邦教育科研部、及汉堡卓越研究群体等大型科研合作项目首席科学家。获多项国内外奖励及荣誉称号,包括中国国家(海外)杰出青年基金(2001年)、2008年被选为美国医药和生物工程学会(AIMBE)会士(Fellow)、2020年当选德国国家工程院院士。
